106
SELECTION TYPE THEORIES*
LINDLEY DARDEN AND JOSEPH A. CAIN
†Committee on the History and Philosophy of Science
University of Maryland, College Park
Selection type theories solve adaptation problems. Natural selection, clonal selection for antibody production, and selective theories of higher brain function are examples. An abstract characterization of typical selection processes is generated by analyzing and extending previous work on the nature of natural selection. Once constructed, this abstraction provides a useful tool for analyzing the nature of other selection theories and may be of use in new instances of theory construction. This suggests the potential fruitfulness of research to find other theory types and construct their abstractions.
1. Introduction. This paper discusses abstract characterizations of selection theories. Finding such abstractions is a task in a larger research program, which is based on the assumption that some scientific theories are representative of types of theories that solve types of problems. Natural selection, clonal selection for antibody production, and selective theories of higher brain function are examples of selection type theories. Selection theories solve adaptation problems by specifying a process through which one thing comes to be adapted to another thing.
When Darwin elaborated his theory of natural selection in 1859, he provided a new type of theory for explaining adaptation problems. Others, such as Burnet (1957) in immunology, have argued by analogy from Darwinian natural selection for selection processes in other fields. One analysis of analogy is that two analogues share a common abstraction (Genesereth 1980). Thus, an analysis of natural selection and its analogues aids in the development of an abstraction for selection theories. The selection literature in philosophy of biology includes several discussions of natural selection that provide help in this task. The abstraction for natural selection extracted from these discussions will then be used in analyzing clonal selection for antibody production in immunology and
*Received September 1987; revised January 1988.
†Lindley Darden thanks the College of Arts and Humanities at the University of Maryland for a Special Research Assignment for support of this work. Thanks also to Michael Bradie, David Casey, Marjorie Grene, Joel Hagen, Pamela Henson, Joshua Lederberg, Elisabeth Lloyd, Elizabeth Napier, James Platt, Frederick Suppe, and two anonymous referees for their comments and ideas.
Philosophy of Science, 56 (1989) pp. 106-129.
Copyright 1989 by the Philosophy of Science Association
107
SELECTION TYPE THEORIES
selective theories for higher brain function in neurobiology.1 Before beginning the analysis of selection theories, however, we will briefly discuss the concept of an abstraction.
2. The Concept of an Abstraction. An abstraction provides a schematic outline that can be filled in or "instantiated" to give an actual theory. It delineates typical components of theories of a given type. Although little work has been done in history and philosophy of science on abstract types of theories, some is suggestive.
Hanson, (1961) in discussing reasoning in discovery, suggests that one might reason that problematic phenomena can be explained by a "type" of hypothesis. Hanson says little about what constitutes a type of hypothesis, but he gives an example: Newton’s theory of gravitation is an inverse-square type of theory. Despite its lack of development, Hanson’s idea has appeal: find types of hypotheses proposed in the past and analyze the nature of the puzzling phenomena to which the type applied. Use this knowledge of a type of hypothesis in future instances of theory construction. In Darden (1987) this method of theory construction is referred to as using the history of science for finding "compiled hindsight".
Suppe (1979) suggests that investigations of types of theories is an important research task in philosophy of science and mentions two types that Shapere (1974, 1977) discusses: compositional and evolutionary. Compositional theories are theories that explain by appealing to component parts, such as atomic theory. Evolutionary theories claim that change over time has occurred, such as biological evolution, stellar evolution, and theories concerning the evolution of the chemical elements. It is important to distinguish selection theories, which we will discuss, from evolutionary theories, on which Shapere focuses. Darwinian natural selection is one explanation for biological evolution, but other theories have been proposed to explain it, such as inheritance of acquired characters and orthogenesis (development toward an end driven by some sort of internal dynamic without regard to environmental conditions). Furthermore, selection theories, such as the clonal theory of antibody production, can also be used to explain adaptive processes that are not evolutionary.
Working at a less abstract level, Kitcher (1981) suggests that theories
1
A note about terminology is useful at the outset. In using the term "natural selection" in contrast to clonal selection and neuronal selection, we don’t mean to imply that the latter two are examples of "artificial" selection. We are following accepted usage in restricting natural selection to the theory that explains adaptations arising through evolutionary time. Although the immune system and brain evolved, the theories about them under discussion explain adaptations within a single individual during its life to combat disease or to learn from experience. Also, we are using "adaptation" in a more general way than the restricted sense of the product formed as a result of natural selection; see, for example, Gould and Vrba (1982).
108
LINDLEY DARDEN AND JOSEPH A. CAIN
can be viewed as providing schematic argument patterns. He develops instructions for instantiating such schemas. His examples include Newtonian laws and a pattern provided by Darwin for explaining the evolution of a particular species. Kitcher’s "schemas" are less abstract than the level at which we will discuss selection type theories. His cases show the use of a single schema within one domain to explain numerous, similar instances. Thus, for Kitcher, using the same schema within a single domain provides a kind of explanatory unification for that area of science. Our discussion of selection theories will be for analogous theories found in separate fields. Use of the same type of theory merely shows that similarly structured theories can be used to explain different domains; it does not unify the areas in the strong sense discussed by Kitcher. However, if one has a realistic interpretation of theories, then finding the same abstract type of theory in numerous domains may be taken as an indication that nature uses similar processes in diverse phenomena. Such a conclusion might be regarded as a weaker form of unity in science than that discussed by Kitcher.
Brandon (1980) also uses the language of "schema" and "instantiation." In discussing the principle of natural selection, he regards the principle of natural selection as a "schematic law". The schema may be "cashed out" in terms of differences in particular traits that apply "to particular populations under particular environmental conditions" (Brandon 1980, pp. 432-433).
Despite this previous work, historians and philosophers of science have not taken as a primary task finding types of problems and types of theories to solve them.2 To find other previous work that seeks theory types and attempts to construct abstractions of them, we must turn to another field.
2
The question may be raised of how our analysis relates to axiomatic (for example. Williams 1970) and semantic (for example. Beatty 1980; Thompson 1983; Lloyd 1984) analyses of evolutionary theory. Our analysis is compatible with these approaches, but has different motivations and aims. Darden was motivated to undertake this analysis as a result of her work on analogy and theory construction (Darden 1982, 1983). The aim here is not to use or provide a general analysis of the structure of all scientific theories. It is an open question whether all theories can be used to form abstractions; furthermore, it is likely that some theories are unique and not examples of recurring types. Instead, the aim is to provide one theory formation strategy, namely, evoking a theory type. Numerous other strategies for theory construction exist (see, for example, Darden 1980, 1982, and 1986). Thc idea of instantiating variables in different ways to produce different theories also has some similarity to Schaffner’s (1980) analysis of theories of the middle range as a series of overlapping, temporal models. Clonal selection is one of Schaffner’s examples, although he does not focus on its nature as a selection type theory. None of these previous analyses, however, shares with ours the desire to construct abstractions that capture structure shared by analogous theories in different fields. Nonetheless, analyzing theories as abstract schemas or models that can be instantiated in different ways may be a fruitful idea to pursue because it has been arrived at from quite different perspectives by a number of philosophers.
109
SELECTION TYPE THEORIES
Recent work on reasoning by analogy and attempts to implement analogical reasoning in computer systems using techniques in artificial intelligence (AI) have produced additional examples of abstract theory types. Gentner (1983) discusses central force systems, such as the solar system and the Bohr model of the atom. Greiner (1985) constructs an abstraction for flow problems, for example, water and electric flow. Thagard and Holyoak (1985) investigate wave type theories, including water waves and sound waves.
The analysis of two analogous things as sharing a common abstract structure (Genesereth 1980) has proved to be fruitful in AI. Methods for forming abstractions and for instantiating them are an active area of research, not only in the analogy work but in research on problem-solving more generally.3 Philosophers, insofar as they have considered abstractions, have tended to think of one level of abstraction of a particular theory, such as a formal set of equations embodied in the theory. But methods of knowledge representation and of implementing reasoning strategies in AI have led to the idea of multiple levels of abstraction (Korf 1985).
Imagine a scientist confronted with the problem of explaining how one thing comes to be adapted to another thing–how an immune system comes to be adapted to fighting an invading antigen or how a brain becomes adapted to responding to a type of signal. History of science provides the compiled hindsight that selection theories have been useful in solving adaptation problems (Darden 1982, 1987). So, our imaginary scientist asks: What should be the case in the problem situation if a selection theory is to provide a useful solution? What needs to be specified in order to construct a selection theory? In order to answer this scientist, our task is to construct an abstraction for selection theories. By instantiating the abstraction, our imaginary scientist may be able to generate testable hypotheses.
Several methods for forming abstractions exist. An abstraction can be constructed from one example by replacing constants with variables. Alternatively, more than one example with the same structure can be used, as in analogical reasoning. Abstracting involves pruning away content and leaving essentials (or "typicals," if one isn't looking for necessary conditions). Comparison of two examples is a good way to locate what is most important. Another way of forming abstractions is to consider several discussions of abstract components of a theory and put complementary parts together in a piecemeal way. (For additional methods, see Darden 1987.)
3
For an algorithmic approach to adaptation see Holland (1975). For a comparison of Holland’s adaptive theories and selection theories, see Darden (1983).110
LINDLEY DARDEN AND JOSEPH A. CAIN
Our task now is to proceed to develop an abstraction for selection type theories. We will draw on recent discussions in the philosophy of biology literature to develop an abstraction for natural selection. The construction of that abstraction will be constrained by our desire to make it applicable, possibly at a slightly higher level of abstraction, to clonal selection and selective theories of brain function. After stating the abstraction for natural selection, we will instantiate it in the other two analogous cases.
3. The Process of Natural Selection. A selection process may be broken down into a series of steps from which a more abstract characterization can be developed.
(A) First are the preconditions before a selective interaction. These include a set of individuals that vary among themselves. Also, the individuals must be in an environment with critical factors that provide a context for the ensuing interaction.
(B) The actual step of selection involves an interaction between individuals and their environment. Because they vary, different individuals will interact differently.
(C) Several types of effects result from the differential interactions. In the short-range, individuals benefit or suffer. If the individuals can be located in a hierarchy (such as gene, organism, group), then there may also be short-range effects of sorting at other levels.
(D) Longer-range effects may follow the short-range effects of the interaction, such as increased reproduction of individuals with certain variations or reproduction of something associated with those individuals.
(E) Even longer-range effects may also occur, such as accumulation of benefits through numerous generations to produce a lineage of individuals.
Our task now is to discuss these steps in more detail and develop an abstraction with appropriate variables.
A. Preconditions for Natural Selection. Natural selection operates on a population of varying individuals in a given location, such as the classic example of the peppered moths in a region with trees darkened by industrial pollution. This case illustrates the following preconditions for selection: the set of individuals making up a population, the presence of the variation, and the environment. Although variation is often discussed as a precondition for selection (for example, Lewontin 1970; Sober 1984), the other preconditions that we list have received less attention. In addition to varying, the individuals
111
SELECTION TYPE THEORIES
on which selection acts must share some features. If the population is an interbreeding population, then the organisms, of course, share many genes and their associated characteristics. We will assume that a set of individuals is delineated by specifying features that they share, not by enumerating its members. Thus our abstraction has as a first precondition that a set of individuals exists. "Group" in Hull’s (1980, 1981) sense might be a better word that "set"; groups tend to be spatiotemporally localized entities and their members are considered part of the group because of their location (Hull 1980, p. 314). But using "group" would introduce confusion with the different issue of whether "group selection" exists. The term "class" might avoid introducing perhaps extraneous connotations from set theory, but Kitcher (1987, p. 186) argues that it introduces confusions of its own.
Hull stresses that selection "can act only on spatiotemporally localized entities" (Hull 1980, p. 314). Sober makes a related claim when he argues that a "common causal influence" operates in a selective process:
Think of two populations living on opposite sides of the universe, each driven by its own internal dynamic. Suppose that one group ends up outreproducing the other. If the two populations are not subject to a common causal influence, then it makes no sense to view them as participating in the same selection process. (Sober 1984, p. 274, footnote 40)
In order to make more explicit the idea, which is perhaps implicit in the views of Hull and Sober, we will state that the set of individuals must be "in an environment E.
" This phrase has the advantage of allowing for further elaboration of the nature of the environment, as we will shortly see.Much can be said about the nature of variation within the set. Sober (1984) reduces his discussion of selection to the simplest case: individuals either have or do not have a single variant property P. Darden (1982) uses the phrase "array of variants", which implies a range of variations within a set. She also discusses the scale of the variants relevant to natural selection. The amount of difference between variants has been a matter of dispute historically; Darwin’s small scale individual differences and de Vries’s (l903-04) large scale mutations are examples. Another issue that may arise within a discussion of the preconditions or selection is how the variants are produced. But as long as preexisting diversity exists, the details of the mechanism of its production can be omitted from a selection type theory.
A key precondition for selection that has received comparatively little discussion in the philosophy of biology is the nature of the environment. (Brandon in an as yet unpublished work has recognized this omission
112
LINDLEY DARDEN AND JOSEPH A. CAIN
too.) Cain (unpublished) stresses the role that critical environmental factors play in creating conditions for a selective interaction. The components of the environment that affect the nature of the struggle for existence will he referred to as "critical factors". What factors in the environment are critical determines what variant properties are relevant to surviving (or otherwise benefiting from) the selective interaction. Thus, the fit between variant and environment (or in a finer-grained analysis, the fit between the variant property and the critical factor) is the key aspect of the next step, the selective interaction.
B. Selective Interaction. Much of the early discussion in philosophy of biology about the nature of selection focuses on the effects of selection, namely differential survival and reproduction. If no account can be given of selection except in terms of the effects, then the specter of tautology is raised: "survival of the fittest" becomes "survival of those that survive".
Lewontin, in his now classic 1970 article discussing the steps in selection and the properties of evolving entities, states: "different phenotypes have different rates of survival and reproduction in different environments" (Lewontin 1970, p. 1). But, we may ask, why is reproduction differential? Lewontin lists three properties of entities that evolve: variation, reproduction and heritability. A fourth component, one involving interaction during the selection step, must be added to this list in order to explain how variation gives rise to differential reproduction. In an earlier attempt at producing an abstraction for natural selection, Darden characterizes the selection step common to artificial selection and Darwinian natural selection. She writes: selection of a subset of variants occurs by an agent selecting according to a criterion (Darden 1982, p. 16). But "agent" brings inappropriate negative analogy from the artificial to the natural case.
A better characterization of the selection step can be constructed using ideas from Hull (1980) and Sober (1984). Especially important are the ideas of interaction and the causal role of the variant property. Their ideas have to be augmented by more detail about the role of the critical factor in the environment. Hull recognizes the nexus of the selective event:
Reproduction by itself is sufficient for evolution of sorts, but not evolution through natural selection. In addition, certain entities must interact causally with their environments in such a way as to bias their distribution in later generations. (Hull 1980, p. 317)
Sober (1984) stresses the causal nature of the selective interaction even more than Hull did. Sober advances us on our way to producing an abstraction for natural selection by providing somewhat helpful character-
113
SELECTION TYPE THEORIES
izations of group and genic selection:
[There] is group selection for groups that have some property P if (and only if)
1) Groups vary with respect to whether they have P, and
2) There is some common causal influence on those groups that make it the case that
3) Being in a group that has P is a positive causal factor in the survival and reproduction of organisms.
The analysis of genic selection goes in parallel. There is selection for possessing the gene P if (and only if)
1) Organisms vary with respect to whether they have P, and
2) There is a common causal influence on those organisms that makes it the case that
3) Possessing the gene P is a positive causal factor in the survival and reproduction of organisms. (Sober 1984, p. 280)
Sober stresses the relation between the possession of a property and the causal role it plays in producing a benefit. Sober provides neither a similar analysis for organismic selection nor an abstraction for natural selection that is independent of hierarchical levels. We may attempt to put variables into Sober’s accounts:
There is X selection for Y's that have some X-level property P if (and only if)
1) Y's vary with respect to whether they have P
2) There is some common causal influence on Y's that makes it the case that
3) Y's possession of P is a positive causal factor in the survival and reproduction of Z's.
X is the hierarchical level considered as the level of selection. Y's are the objects that vary. Z is the level that Sober calls the "benchmark". A benchmark is an object that benefits or suffers in virtue of having properties that are selected for (Sober 1984, p. 270). However, the relation of Y to X and Z is unclear. For the genic selection example Y = Z = organisms. But in the group selection case, Y is not equivalent to Z: Y = groups, but Z = organisms. Y's are the variants, but Sober is unclear about whether the variation is to be located at the level of causal influence (Y) or at the benchmark (Z). For simplicity, initially we are going to assume that the variant, the interactor, and that which directly benefits or suffers can be represented by the same variable. Alternative, lower-
114
LINDLEY DARDEN AND JOSEPH A. CAIN
order abstractions can be constructed that differentiate these three variables. Sober’s analysis introduces complexities because he is grappling with the issue of effects at various hierarchical levels. Since Lewontin’s (1970) analysis of selection at different levels, others have also tried to analyze selection and its effects at different levels in structural or functional hierarchies (for example, Hull 1980: Arnold and Fristrup 1982). In the abstraction we are constructing, we wish to avoid most of the complexity the hierarchical perspective on selection introduces. However, we will use one distinction used by Vrba and Gould (which parallels a similar distinction in Sober): sorting and selecting.
In Darwinian theory, evolutionary change is the product of sorting (differential birth and death among varying organisms within a population). Sorting is a simple description of differential representation; it contains, in itself no statement about causes. As its core, Darwinism provides a theory for the causes of sorting–natural selection acting upon organisms in the "struggle for existence." However other processes (genetic drift, for example) produce sorting as well; thus, the two notions–sorting and selection (a favored theory for the cause of sorting)–are quite distinct and should be carefully separated. (Vrba and Gould 1986, p. 217)
Vrba and Gould’s definition of selection makes use of their distinction between sorting and selecting and also shows their localization of the selective interaction at the hierarchical level of the variant property:
We suggest that individuals are subject to selection at any level for the characters they possess only if these characters are heritable and emergent, and if they interact with the environment to cause sorting . . .: Selection encompasses those interactions between heritable emergent character variation and the environment that cause differences in rates of birth or death among varying individuals. (Vrba and Gould 1986, p. 219; emphasis theirs)
The single most important omission from the previous discussions is a neglect of the role that environments play in selective interactions. Although Hull and Vrba and Gould allude to the interactions with the environment, no analysis is given of conditions in an environment that are relevant to the selective interaction. Sober doesn’t explicitly mention the role of the environment in his schematic discussions of group and genic selection (Sober 1984, p. 280). Consideration of his steps shows that he packs the role of the environment into "common causal influence" and into his analysis of how a variant property can be a "positive causal factor" in selection. We analyze the environment in terms of the critical
115
SELECTION TYPE THEORIES
factor that makes a given property causally relevant to the selective interaction. The critical environmental factor and the interaction of the variant property with it are the keys to the subsequent effect of the selective interaction.
In summary, the main points about the selective interaction are the following: first, an explicit recognition that a step in the temporal process occurs between the formation of variants and the effect of differential reproduction; second, that this step involves interaction; third, the interaction can be analyzed either in terms of an individual and its environment or, in a finer-grained way, in terms of the variant property and the critical factor in the environment; fourth, the variant property plays a causal role in the interaction; and, finally, effects can be manifested at levels other than the level of the interaction.
C. Effect. Sober uses the language of "benefit and suffer" to characterize the differential effects of the selective interaction. Although such language sounds somewhat anthropomorphic and will require a value judgment as to what is a benefit, it is appropriately abstract for our purposes for a number of reasons. In natural selection, the interaction may produce an effect that only later results in differential survival and reproduction. Thus, the process should have a step for specifying such an intermediate effect. Also, the benefits in the many different instances of selection events will be of many different types, for example, escaping predators, obtaining food, defending territory. "Benefit" is a neutral term for characterizing numerous types of positive effects.
Most importantly, the neutral language of "benefit" is desirable for constructing an abstraction that can apply at different levels in an organizational hierarchy within an evolutionary theory context. It is even more desirable for our current purpose of constructing an abstraction that can be instantiated in other selection type theories. We would be happy with less anthropomorphic and value-laden sounding term than "benefit", but we haven’t yet found one at an appropriately abstract level.
D. and E. Longer-Range Effects. We have carefully delayed adding differential reproduction to our steps in a selection process until these later steps. As others have argued, defining selection as differential reproduction masks many of the important features of selection. For artificial selection, it may be very important to separate the benefit from differential reproduction. The breeder might select on a criterion of low seed output in grapes; hence, decreased reproductive capacity actually results in more grape plants being grown. Nonetheless, for natural selection, differential reproduction as a result of the differential interaction is the important consequence that makes a difference in evolutionary change.
116
LINDLEY DARDEN AND JOSEPH A. CAIN
Differential replication is an integral part of Hull’s analysis of selection, as is the even longer-range consequence of the production of a lineage. Hull defines a lineage as, "an entity that changes indefinitely through time as a result of replication and interaction" (Hull 1980. p. 327).
We have discussed steps A-E and will turn to a construction of a more abstract characterization of them, attempting to be more precise and use variables in place of many of the words of the looser preliminary list.
4. Abstraction for Natural Selection. In considering what aspects of the various discussions should be preserved in an abstraction for selection theories, the following components are important. Darden's idea of an "array of variants" focuses attention on the fact that one needs to specify a set of things that vary. Implicit in that idea is that the individuals in the set share many properties but vary in one or more. For simplicity, Sober suggests a single variant property P. Hull stresses causal interaction. Sober's primary emphasis is on the "common causal influence" operating in the selective event and the role of the variant property as a "positive causal factor". He provides appropriate neutral language of the effect being some sort of "benefit". Vrba and Gould provide the distinction between selection at a given level in a hierarchy and sorting at other levels. To these discussions we have added the concept of a critical factor in the environment.
Drawing on this previous work, we now will attempt to develop our abstraction. A typical selection process can be analyzed into a group of preconditions, such that if they obtain, then a selective interaction occurs, resulting in a series of effects. A selection process typically occurs if:
A. Preconditions
i. A set of Y's exists and
ii. Y's vary as to whether they have property P and
iii. Y's are in an environment E with critical factor F.
B. Interaction
iv. Y's, in virtue of possessing or not possessing P, interact differently with environment E and
v. critical factor F affects the interaction such that
vi. the possession of P causes Y's with P to benefit and those without P to suffer.
117
SELECTION TYPE THEORIES
(vi’. This causal interaction may have the concomitant effect of sorting Z's.)
D. Longer-range effect
vii. C may be followed by increased reproduction of Y
’s with P or reproduction of something associated withY’s.
E. Even longer-range effect
viii. D may be followed by longer-range benefits.
To give some substance to these abstract statements, let’s consider the classic case of the peppered moths. (i) A population or set of peppered moths exists. (ii) The moths vary in the property of color, dark or light. (iii) Moths are preyed upon by birds and live in woods near coal-burning plants where trees are darkened by soot. (iv) Moths, in virtue of possessing or not possessing dark color, interact differently with birds. (v) The darkness of the trees affects the interaction such that (vi) the possession of dark color causes moths with dark color to benefit and those without dark color to suffer. (This causal interaction may have the concomitant effect of sorting genes in the moth population.) (vii) Those moths with dark color tend to produce more offspring, which inherit dark color.
The abstraction is a simplification in a number of explicit ways. In condition (i) the existence of a set is specified, but common properties that define the set are not explicitly given, though they could be. Condition (ii) is the simplest case in which Y's differ by only one property. If the way in which Y's differ is unknown, or Y's differ in more than one property, this condition could be altered accordingly. Condition (ii) considers the range of the variant property, which here is stated in the simplest possible way, that is, having or not having P, could be altered to reflect other possibilities, such as a continuous gradation of some sort or quantitative differences. (See Darden (1982) for a discussion of instantiating details about variation.)
Specifying an environment and a single factor within it that is relevant to the selective interaction (conditions iv and v) may sometimes be difficult. How to delineate the relevant "environment" from everything else may be a formidable task. Limiting discussion to a single environmental factor F is a simplification. In the moth example, the environment includes the nature of the predator as well as environmental conditions, for example, color of trees. Thus sometimes the effects of the critical factor are mediated through an agent, such as a predator.
Condition (iv) of interaction is separated from the causal condition (vi) because it is possible to have different interactions without having dif-
118
LINDLEY DARDEN AND JOSEPH A. CAIN
ferential benefits. For example, if toads have tongues of different lengths, then those with longer tongues can catch insects at a greater distance. But if insects are sufficiently plentiful, then all toads get adequate food despite this differential interaction; no relative benefit or suffering results. Some critical factor in the environment, condition (v), must produce a situation that makes P causally relevant to benefiting.
With some trepidation, we have chosen to follow others and include causal language in our abstraction. We are not here providing an analysis of the "cause" in our abstraction. We are merely stating that in a typical selection process the interaction between variant property P and the environmental factor F causes those variants with P to benefit. Explaining the way differences in interaction cause differences in benefit will depend on case-specific information. We realize the statistical nature of the process and the implicit ceteris paribus clause: sometimes those with P will be in appropriate conditions to benefit, but other times something else will befall them and they won't actually benefit. "Benefit" and "suffer" are vague, but this vagueness has the advantage of allowing instantiation in numerous ways in different specific cases. Benefit could be interpreted as contributing to survival and reproduction (with inheritance of the beneficial property), but it need not. Benefit can be measured instead in terms of immediate effects on Y's. Theories lacking a reproduction step might be called "election" rather than "selection" theories.4
The parenthetical remark in (vi’) about the concomitant effect of a selective interaction sorting Z's makes use of Vrba and Gould's distinction between sorting and selecting. Our abstraction clearly separates Y's that are selected and Z's that are sorted but not directly selected. Whether sorting in this sense actually occurs depends on whether or not there are hierarchical features in the particular case. The variable Y is used in three different ways in our abstraction–in (A) as a variator, in (B) as an interactor and in (C) as the benefitor. In a more fine-grained analysis, one might propose three related, but separate variables, which need not be restricted to a single hierarchical level.
5. Clonal Election Theory for Antibody Formation. The goal of this work is to develop an abstraction for selection type theories. Thus far, we have developed an abstraction for natural selection. Now we will consider another successful selection theory, which comes from immunology. In 1955, Jerne proposed the natural selection theory for antibody
4This terminology was used in Lederberg (1959). Shapere and Edelman in Ayala and Dobzhansky (1974, pp. 202-204) explicitly separated reproduction and differential reproduction in their "criteria of natural selection",
119
SELECTION TYPE THEORIES
formation. Jerne’s theory was subsequently modified by Burnet (1957) to the theory of clonal selection.
The problem is to explain how the immune system forms antibodies that are able to deactivate many different types of foreign substances (called antigens) that invade the body, while not attacking the body’s own substances. Jerne, in reflecting on the components of his theory, says:
three mechanisms must be assumed: (1) a random mechanism for ensuring the limited synthesis of antibody molecules possessing all possible combining sites, in the absence of antigen, (2) a purging mechanism for repressing the synthesis of such antibody molecules that happen to fit auto-antigens, and (3) a selective mechanism for promoting the synthesis of those antibody molecules that make the best fit to any antigen entering the animal. (Jerne 1966, p. 301)
Thus Jerne proposes several mechanisms: one for producing variants, another for negative selection against any antibodies that would attack the body’s own substances, and a third positive selective mechanism for increasing production of antibodies against an invading antigen. The positive selective mechanism works, according to Jerne, by a specific, circulating antibody attaching to an invading antigen, the complex of antigen-antibody being engulfed by a cell, and that cell (or perhaps another one that is signaled) beginning to produce more of the specific antibody molecule. Jerne, writing before the details of protein synthesis had been worked out in molecular biology, admits that the idea that a protein molecule entering a cell could signal the cell to produce more molecules of that kind is an "unfamiliar" notion (Jerne 1955, pp. 849-850).
Although Burnet (1957) endorses much of Jerne’s theory, he objects to the mechanism of production of antibodies and proposes an alternative at the level of the cell rather than the molecule. A similar change in level from molecule to cell was proposed (apparently independently) by Talmadge (1957). (For further discussion of Talmadge’s role see Burnet (1957, p. 67); Talmadge (1986, p. 7); and Ada and Nossal (1987, p. 65). Jerne (1966, p. 308) in a retrospective on the theory in 1966 gives credit to Burnet and does not mention Talmadge.)
Of Jerne’s theory, Burnet says:
Its major objection is the absence of any precedent for, and the intrinsic unlikelihood of the suggestion, that a molecule of partially denatured antibody could stimulate a cell, into which it had been taken, to produce a series of replicas of the molecule.... it would be more satisfactory if the replicating elements essential to any such theory were cellular in character ab initio rather than extracellular
120
LINDLEY DARDEN AND JOSEPH A. CAIN
protein which can replicate only when taken into an appropriate cell. (Burnet 1957, p. 67)
Thus Burnet changes the level of organization of the unit that replicates from molecule to cell. Furthermore, he claims that the cell is the unit of variation; one cell produces one type of antibody. He proposes somatic mutation as the mechanism of the production of variants. In addition to making the cell the unit of variation and of reproduction, he also makes it the unit of interaction; the lymphocyte cell has a reactive site that is "equivalent" to the antibody it produces. The reaction between the reactive site and antigen serves as a signal to the cell to produce a clone of cells like itself. The cloned cells then release tree antibodies of the specific type to attack the invading antigen. (For additional discussion, see Lederberg 1959; and Schaffner 1980, pp. 69-71.)
Jerne and Burnet’s theories illustrate additional instances of selection type theories that may be analyzed to yield an abstraction. The differences in their theories show that the variables can be instantiated in different ways either to improve a theory or to yield a competing alternative. In this case, the level of organization–molecule or cell–provides an important point of difference in the ways the variables are instantiated.
Jerne’s and Burnet’s theories attempt to solve an adaptation problem: how is the immune system’s defense against an invading antigen produced, or, more specifically, how are antibodies against an antigen formed? Fighting invaders is an adaptive response of the immune system.
By comparing Burnet’s theory to our previous abstraction, we can develop an abstraction for Burnet’s clonal selection theory.
A selection process occurs if:
A. Preconditions
i. A set of lymphocyte cells exist and
ii. these cells vary as to the reactive sites on their cell surfaces.
iii. These cells are in the blood stream with invading antigens.
B. Interaction
iv. The cells, in virtue of possessing or not possessing certain reactive sites, interact differently with the antigens and
v. the properties of the antigen affect the interaction such that
C. Effect
vi. the possession of a particular reactive site causes cells with such a site to be activated and those without such a site not to be activated .
121
SELECTION TYPE THEORIES
D. Longer-range effect
vii. Activated cells proliferate in clones thereby producing more cells of their type; the cloned cells release free antibodies of a given type that attack the antigen.
E. Even longer-range effect
viii. More cells of that type will be present even after invading antigen has been eliminated and the system returns to "normal"; thus, the immune system has acquired the ability to respond more quickly in the future if an antigen of that type invades.
The selection event involves a differential activation. The effect is a differential amplification of cells that has the concomitant effect of producing more antibodies of the given type.
6. Selection Theories for Brain Function. A number of rather speculative theories for explaining various aspects of brain functioning have been proposed that are selection type theories. These include theories by Conrad (1976); Changeux (1985); and Edelman (1978). Here, we will focus on Edelman’s theory. The adaptation problem is to explain increasing efficiency of the brain in processing a given type of signal. First, how does the brain accommodate new types of signals that have never before been encountered? Second, how does the brain refine a response pattern when it encounters a type of signal repeatedly? (Edelman 1978, p. 55.) Edelman proposes a theory to solve this adaptation problem; he calls it the "group-selective theory of higher brain function":
The basic idea is that the brain is a selective system that processes sensory motor information through the temporally coordinated interactions of collections or repertoires of functionally equivalent units, each consisting of a small group of neurons [brain cells]. (Edelman 1978, p. 52; emphasis his)
The units of variation are groups of neurons. Groups having nearly identical functions are part of a repertoire, in Edelman’s terminology (p. 62), or a set of variants, in our terminology. Functionality of groups of neurons is defined by their response to signals. Although all the groups in a repertoire share the property of being able to respond to a given signal, these groups vary in their internal structural organization. In Edelman’s terminology (and used similarly in molecular biology) the groups are degenerate, not redundant: groups with different intrinsic connections respond to identical stimuli. "Degenerate groups are isofunctional, but
122
LINDLEY DARDEN AND JOSEPH A. CAIN
nonisomorphic . . . redundant groups are isofunctional and isomorphic" (Edelman 1978, p. 59, Fig. 3).
The variant groups are produced as a result of both genetic factors and "epigenetic" factors, which occur during the embryological and early development of the brain. Such factors include "selective stabilization" of synapses (connections between neurons) discussed by Changeux (Edelman 1978, pp. 88-89). But even after these selection events, which give rise to structurally different neuron groups, much degeneracy remains, according to Edelman. This degeneracy provides the variability for the selective interactions that occur between the neuron groups and the signal input throughout the life of the organism.
By some means that Edelman does not specify, the variation in internal structure of each particular group determines how well it will respond to a given stimulus. As a result of the variation in structure, some neuron groups are more easily stimulated or more often stimulated than others. That stimulation has the effect of either reinforcing a particular neuron group or somehow selecting against the other groups in that repertoire, thus altering their potential for subsequent stimulation by similar signals. The accumulation of such alterations has the longer-range effect of building up a secondary repertoire of groups. In other words, the reinforced groups have the potential to respond more efficiently in the future.
Edelman’s work is obviously only in the early stages of theory construction. Many details remain to be specified. Constructing an abstraction following the pattern from the natural selection and clonal selection theories is a method that Edelman himself recognizes as valuable. (See his Table 1: "Some Characteristics of Selective Systems", Edelman 1978, p. 91.) The following is our attempt at instantiating our abstraction with Edelman’s theory.
A selection process occurs if:
A. Preconditions
i. A set of degenerate neuron groups exist that respond to a given signal (that is, an isofunctional repertoire
exists), and
ii. neuron groups vary in intrinsic connections and thus in their abilities to respond to a given signal, and
iii. neuron groups are in regions of the brain that receive a given sensory signal with a particular spatiotemporal
configuration such that
B. Interaction
iv. neuron groups, in virtue of their different intrinsic connections, interact differently with the sensory signal,
and
123
SELECTION TYPE THEORIES
v. the particular spatiotemporal configuration of the signal affects the interaction such that
C. Effect
vi. the possession of certain intrinsic connections causes neuron groups with such connections to be stimulated
and/or those without such connections to he inhibited.
(vi’. This causal interaction has the concomitant effect of sorting individual neurons.)
D. Longer-range effect
vii. Stimulation of neuron groups causes reinforcement of those groups.
E. Even longer-range effect
viii. A secondary repertoire of committed neuron groups develops for responding more efficiently to signals
similar to those previously encountered.
Instantiating the variables in our abstraction for Edelman’s theory shows the vagueness in his theory as to the nature of the relation between the variations and the critical factor in the signal that interacts with the variant groups. In other words, what is it about the intrinsic connections that causes some groups to have a better "match" (Edelman 1978, p. 56) to a signal? In terms of our variables, the Y
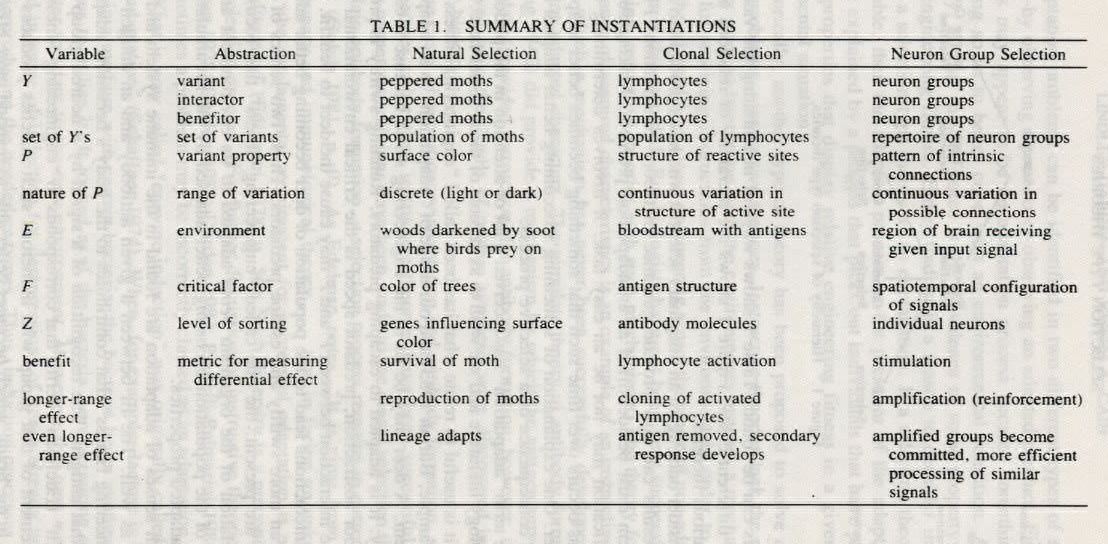
’s have in common the ability to process a given signal, but Edelman’s analysis has not yet provided values to instantiate P and F; more specifically, what property of the intrinsic connections (P) causes them to process a given signal configuration (F) in a better or worse way? Edelman tells us too little about the relation between neuron connections and signal configurations to allow us to see why some would "fit" better than others. Our abstraction suggests further theory construction needs to be done on this point.Table I shows the instantiations for the variables in the abstraction extracted from the three cases. It shows the analogies between the three theories and the components of Edelman’s theory that we have used in instantiating his theory in our abstraction.
7. Conclusion. The abstraction extracted from the characterizations for natural selection proved of sufficiently high order for the immunology and neuronal cases, except that the longer-range effects weren’t necessarily reproduction. A disjunction, such as "reproduction or amplification or reinforcement", might serve unless some more abstract term for these
124
125
SELECTION TYPE THEORIES
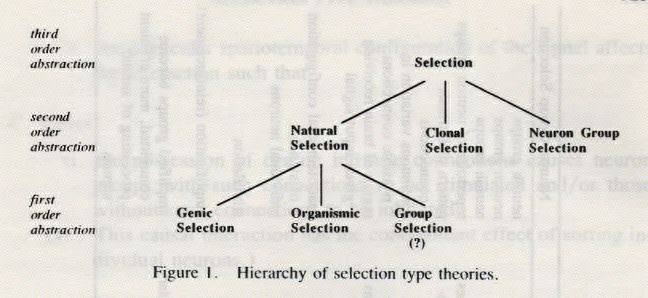
payoffs can be found. Table 1 shows ways of instantiating the variables in the different cases. Figure 1 shows the hierarchy of relations among the various theories.
Consider once again an imaginary scientist who has a problem to solve. Our results can now provide guidance in theory construction. First, a problem must be identified as an adaptation problem. Making such an identification may not be an easy task. It involves determining that a process occurs over time such that something becomes better adapted to another. One explanation of something’s becoming better adapted is that it is a consequence of a selective process.
The term "adaptation" has received a lot of attention in the philosophy of biology literature and we wish to skirt most of those issues here. However, it is useful to relate what has been called "engineering fitness" (see for example, Burian 1983) and our usage of "adaptation" in an adaptation problem. A deer may be fit in the engineering sense because it can run swiftly: it is adapted to escaping predators. Running swiftly is property P in our abstraction; predator speed is the critical environmental factor F. The interaction between P and F may cause the deer to benefit. Many such interactions lead to the population of deer becoming better adapted: they run more swiftly and escape predators. Thus, two senses of adaptation are present: the one called "engineering fitness" that focuses on the relation of the organism being adapted to its environment and the sense of population becoming better adapted by the accumulation of such adapted properties.
Both senses of adaptation are found in the immune system and brain cases as well. The fit between a given antibody and an antigen is the "engineering fitness" that contributes to the body’s becoming adapted to fight that antigen. The recognition of a signal by a neuron group is the "match" (Edelman’s term) that corresponds to "engineering fitness"; reinforcement of that group with numerous such matches produces adaptation in the brain, which becomes better able to respond more efficiently to such types of signals.
126
LINDLEY DARDEN AND JOSEPH A CAIN
Adaptation problems can be identified in two different types of situations–by observing a process or observing an adapted product. First, a process may appear to be an adaptation process because something is changing so as to better cope with an environmental condition. Alternatively, all that may be noted is the prevalence of some property with good engineering fitness. Either finding is a reason to consider instantiating a selection type theory as an explanation. If a selection theory is applicable, then the adaptive process or the presence of the adaptation will be explained by a process that picks out individuals that have preexisting properties that display relative engineering fitness in a given environmental situation.
Historically, another type of theory has been proposed to solve adaptation problems–instructive theories. Instructive theories lack the set of variants prior to interaction with the environment. Instead, a mechanism exists for receiving instructions from the environment in order to construct the adapted form, in an engineering sense; the waste of non-adapted variants is thus avoided. But biologists have not been very successful in proposing instructive theories with such mechanisms. The theory for the inheritance of acquired characteristics is an example of an instructive theory in which a mechanism has never been successfully developed for acquiring adapted characters. The template theory of antibody formation (Pauling 1940) did provide a mechanism by which the antigen served as a template (provided instruction) for antibody formation; however, it was disproved once the details of protein synthesis were discovered in molecular biology.
In the cases of evolutionary adaptation and antibody formation, selection theories rather than instructive theories have been successful. Although numerous additions have been made to the clonal selection theory to explain antibody formation, a selection process remains in current accounts (Golub 1981, p. 9). As Edelman notes, "It is clear from both evolutionary and immunological theory, that in facing an unknown future, the fundamental requirement for successful adaptation is preexisting diversity" (Edelman 1978, p. 56). Such diversity is a precondition for a selection process. (For further discussion of instructive theories see Edelman 1978; Darden 1982; Darden 1987; Piattelli-Palmarini 1986.)
Sometimes a problem may be incorrectly classed as an adaptation problem What is now called "enzyme induction" was originally called "enzyme adaptation" (Schaffner 1974). When, for example, lactose is introduced in a medium with certain bacteria, the bacteria begin synthesizing enzymes to utilize it. It looked as if the bacteria were acquiring a new character to adapt to the new environment. However, this case was explained in terms of induction (or derepression) of genes already present to begin producing the needed enzymes. Thus, induction processes share
127
SELECTION TYPE THEORIES
features with adaptation processes and must be distinguished from them in order to determine what kinds of theories to evoke. In induction, a very specific preprogrammed response is initiated; it is a less creative process than are selective and instructive processes, which can give rise to new types of adapted forms.
Once an adaptation problem has been identified, then our analysis suggests considering a selection theory as an explanation for it. Instantiating the variables in our abstraction is one way of constructing a selection theory. Table I provides the blanks to be filled and the abstraction in section 4 provides the outline of the theory. Some variables may be instantiated in different ways (for example, at different levels of organization) to produce alternative hypotheses, which can then be tested to confirm or disconfirm them.
Using a selection theory in a new case provides a measure of "plausibility by analogy" (Darden 1976, p. 145). By invoking a type of process that has been confirmed to apply in other biological cases, a measure of plausibility is provided for the new hypothesis–it is like processes known to occur elsewhere. But "plausibility by analogy" does not substitute for direct empirical evidence. Of course, a new theory will require testing against the data in its own domain. Instantiating a type of theory, even a previously very successful type, is no guarantee that a theory will be confirmed in a new case.
This work demonstrates that at least one abstraction for an important type of theory can be constructed. The extent to which other types of problems and types of theories can be found and abstracted is an important area for future work for historians and philosophers of science. Such work holds the promise of providing "compiled hindsight" (Darden 1987) from the history of science, which may prove useful to current researchers in constructing new theories and, perhaps, providing insights into types of processes common in the natural world in which we live.
REFERENCES
Ada, G. L. and Gustav, N. (1987), "The Clonal Selection Theory", Scientific American 257: 62-69.
Arnold, A. J., and Fristrup, K. (1982), "The Theory of Evolution by Natural Selection A Hierarchical Expansion",Paleobiology 8: 113-129.
Ayala, F. J., and Dobzhansky, T. (eds.) (1974), Studies in the Philosophy of Biology. Berkeley: University of California Press.
Beatty, J. (1980), "What's Wrong with the Received View of Evolutionary Theory", in P. D. Asquith and R. N. Giere (eds.), PSA 1980. vol. 2. East Lansing, Michigan:Philosophy of Science Association, pp. 397-426.
Brandon, R. (1980), "A Structural Description of Evolutionary Theory", in P. D. Asquith and R. N. Giere (eds.), PSA 1980. vol. 2. East Lansing, Michigan: Philosophy of Science Association, pp. 427-439.
–
. (unpublished), Adaptation and Environment, in progress.Burian, R. (1983), "Adaptation", in Marjorie Grene (ed.), Dimensions of Darwinism.
128
LINDLEY DARDEN AND JOSEPH A. CAIN
Cambridge: Cambridge University Press, pp. 287-314.
Burnet, F. M. ( l957),"A Modification of Jerne's Theory of Antibody Production Using the Concept of Clonal Selection", The Australian Journal of Science 20: 67-69.
Cain, J. A. (unpublished, 1987), "The Role of the Environment in Selective Interactions", presented at Summer Conference on the History, Philosophy, and Social Studies of Biology, Virginia Polytechnical Institute and Stale University, Blacksburg, Virginia, June 16-20, 1987.
Changeux, J. P. ( 1985), Neuronal Man: The Biology of Mind. New York: Pantheon, Random House.
Conrad, M. (1976), "Complementary Molecular Models of Learning and Memory", BioSystems 8: 119-138.
Darden, L. (1976), "Reasoning in Scientific Change: Charles Darwin, Hugo de Vries, and the Discovery of Segregation", Studies in the History and Philosophy of Science 7: 127- 169.
–
. (l980), "Theory Construction in Genetics", in T. Nickles (ed.),Scientific Discovery: Case Studies. Dordrecht: Reidel, pp. 151-170.–
. (1982), "Artificial Intelligence and Philosophy of Science: Reasoning by Analogy in Theory Construction", in T. Nickles and P. Asquith (eds.), PSA 1982, vol. 2. East Lansing, Michigan: Philosophy of Science Association, pp. 147-165.–
. (1983), "Reasoning by Analogy in Scientific Theory Construction", in R. S. Michalski (ed.), Proceedings of the l983 International Machine Learning Workshop, Urbana, Illinois: Department of Computer Science, University of Illinois, pp. 32-40.–
. (l986), "Reasoning in Theory Construction: Analogies, Interfield Connections, and Levels of Organization", in P. Weingartner and G. Dorn (eds.), Foundations of Biology. Vienna, Austria: Holder-Picher-Tempsky, pp. 99-107.–
. (1987), "Viewing the History of Science As Compiled Hindsight", AI Magazine 8 (2): 33-41.Darwin, C. [1859] (l966), On the Origin of Species, A Facsimile of the First Edition Cambridge, Mass.: Harvard University Press.
Edelman, G., and Mountcastle, V. (1978), The Mindful Brain: Cortical Organization and the Group Selective Theory of Higher Brain Function. Cambridge, Mass.: MIT Press.
Genesereth, M. (l980),"Metaphors and Models", Proceedings of the First Annual National Conference on Artificial Intelligence. Menlo Park, California: American Association for Artificial Intelligence, pp. 208-211.
Gentner, D. (l983), "Structure Mapping–A Theoretical Framework for Analogy", Cognitive Science 7: 155-170.
Golub, E. (1981). The Cellular Basis of Immune Response, 2nd ed. Sunderland, Mass.: Sinauer Associates.
Gould, S. J., and Vrba, E. (1982), "Exaptation–A Missing Term in the Science of Form", Paleobiology 8: 4-15.
Greiner, R. (1985), "Learning by Understanding Analogies", Ph.D. Dissertation, Department of Computer Science, Stanford University.
Hanson, N.R. (1961), "Is There a Logic of Scientific Discovery?" in H. Feigl and G.Maxwell (eds.), Current Issues in the Philosophy of Science.New York: Holt, Rinehart and Winston. Reprinted in B. Brody (ed.), Readings in the Philosophy of Science. Englewood Cliffs, New Jersey: Prentice-Hall, 1970, pp. 620-637.
Holland, J.H. (1975), Adaptation in Natural and Artificial Systems. Ann Arbor, Michigan: University of Michigan Press.
Hull, D. (1980), "Individuality and Selection", Annual Review of Ecology and Systematics 11: 311-332.
–. (1981), "Units of Evolution: A Metaphysical Essay", in U.L. Jensen and R. Harré (eds.), The Philosophy of Evolution. Brighton: Harvester Press, pp. 23-44. Reprinted in R.N. Brandon and R.M. Burian (eds.), Genes, Populations, and Organisms: Controversies over the Units of Selection. Cambridge Mass.: MIT Press,
1984, pp. 142-160.129
SELECTION TYPE THEORIES
Jerne. N. K.(1955). "The Natural-Selection Theory of Antibody Formation", Proceedings of The National Academy of Sciences 41: 849-857.
–
. (1966), "The Natural Selection Theory of Antibody Formation; Ten Years Later", in J. Cairns, G.S. Stent, and J.D.Watson (eds.), Phage and the Origins of Molecular Biology. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory of Quantitative Biology, pp. 301-312.Kitcher, P. (1981), "Explanatory Unification", Philosophy of Science 48: 507-531.
–.(1987), "Ghostly Whispers: Mayr, Ghiselin, and the ‘Philosophers’ on the Ontological Status of Species", Biology and Philosophy 2: 184-192.
Korf, R.E. (1985), "An Analysis of Abstraction in Problem Solving", in J.J. Pottmyer (ed.), Proceedings of the 24th Annual Technical Symposium. Gaithersburg Maryland: Washington, D.C. Chapter of the ACM, June 20, 1985, pp.7-9.
Lederberg, J. (1959), "Genes and Antibodies", Science 129: 1649-53. Reprinted with a postscript in Stanford Medical Bulletin 19 (1961): 53-61.
Lewontin, R. C. (1970), "The Units of Selection", Annual Review of Ecology and Systematics 1: 1-18.
Lloyd, E. (1984), "A Semantic Approach to the Structure of Population Genetics",Philosophy of Science 51: 242-264.
Pauling, L. (1940), "A Theory of the Structure and Process of Formation of Antibodies", Journal of the American Chemical Society 62: 2643-2657.
Piattelli-Palmarini, M. (1986), "The Rise of Selection Theories: A Case Study and Some Lessons from Immunology", in W. Demopoulos and A. Marras (eds.), Language Learning and Concept Acquisition: Foundational Issues. Norwood, New Jersey: Ablex Publishing Co., pp. 117-130.
Schaffner, K. (1974), "Logic ofDiscovery and Justification in Regulatory Genetics", Studies in the History and Philosophy of Science 4: 349-385.
–
. (1980), "Theory Structure in the Biomedical Sciences", The Journal of Medicine and Philosophy 5: 57-97.Shapere, D. (1974), "On the Relations Between Compositional and Evolutionary Theories", in Ayala and Dobzhansky (eds.) (1974), pp. 187-201.
–. (1977), "Scientific Theories and Their Domains," in F. Suppe (ed.), Structure of Scientific Theories, 2nd ed. Urbana, Illinois: University of Illinois Press, pp. 518-562.Sober, E. (1984), The Nature of Selection. Cambridge, Mass.: MIT Press.
Suppe, F. (1979), "Theory Structure", in P. Asquith and H. Kyburg (eds.), Current Research in Philosophy of Science. East Lansing, Michigan: Philosophy of Science Association, pp. 317-338.
Talmadge, D.W. (1957), "Allergy and Immunology", Annual Review of Medicine 8: 239-256.
–
.(1986), "The Acceptance and Rejection of Immunological Concepts", Annual Review of Immunology 8: 239-256.Thagard, P., and Holyoak, K. (1985), "Discovering the Wave Theory of Sound: Inductive Inference in the Context of Problem Solving", in Proceeding of the Ninth International Joint Conference on Artificial Intelligence. Los Angeles, California, pp. 610-612.
Thompson, P. (1983), "The Structure of Evolutionary Theory: A Semantic Approach", Studies in the History and Philosophy of Science 14: 215-229.
Vrba, E., and Gould, S.J. (1986), "The Hierarchical Expansion of Sorting and Selection: Sorting and Selection Cannot Be Equated", Paleobiology 12: 217-228.
De Vries, H. [1903-04] (1969), The Mutation Theory (2 volumes, translated by J.B. Farmer and A. D. Darbishire, 1909-10). New York: Kraus Reprint Company.
Williams, M. (1970), "Deducing the Consequences of Evolution: A Mathematical Model", Journal of Theoretical Biology, 29:343-385.
{kind=link}
{kind=link}